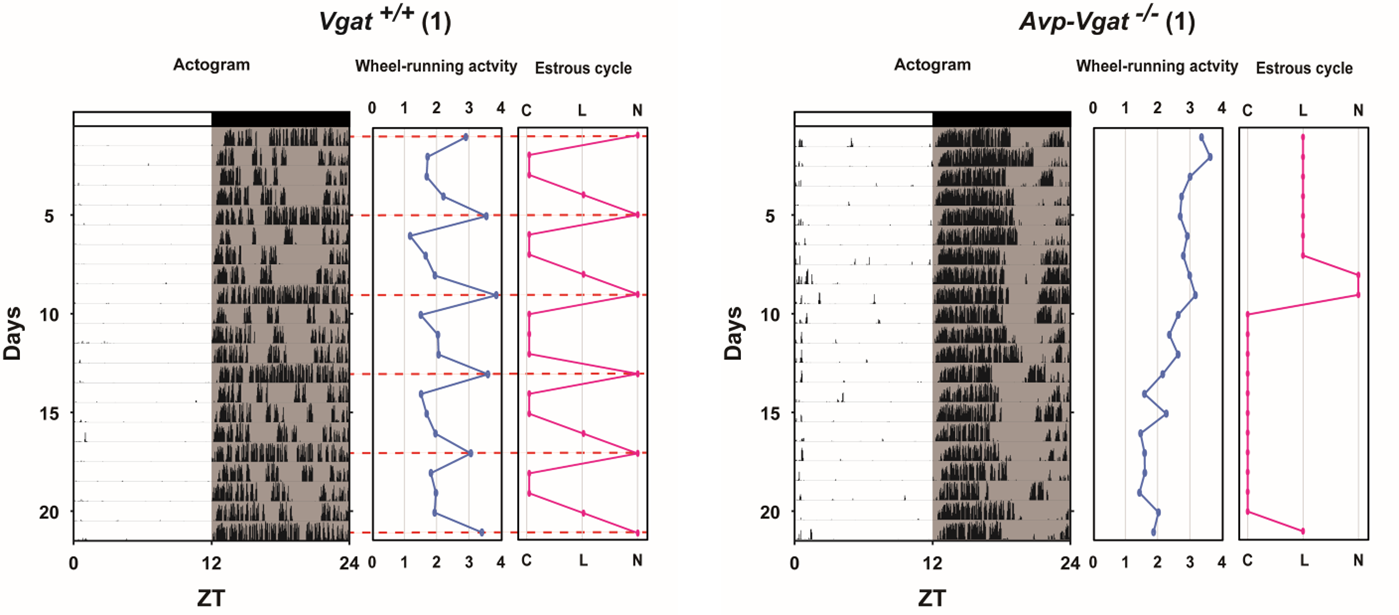
Female reproduction
ヒトを含む多くの雌性動物には、卵巣周期や月経周期といった性周期の存在が確認されており、これは排卵から次の排卵までの期間を指しています。動物実験によく用いられているマウスやラット等の夜行性げっ歯類の性周期は4-5日で回帰しており、発情前期・発情期・発情後期・発情休止期の4ステージに分類されます。性周期に関する生理現象の中で特に重要である「排卵」は、血中エストロゲンの濃度上昇を起点として、性腺刺激ホルモン放出ホルモン(gonadotropin releasing hormone, GnRH)の一過性分泌(GnRHサージ)とそれに続く黄体形成ホルモン(luteinizing hormone, LH)の一過性分泌(LHサージ)により誘発されます(図1)。マウスにおいてこのLHサージは決まって発情前期の夕方に起こる事が知られており、概日時計中枢として知られている脳の視床下部の視交叉上核(suprachiasmatic nucleus, SCN)を破壊すると、規則的なLHサージが観察されなくなり、性周期が乱れる事が報告されています。すなわち、SCNからの伝達情報は排卵性GnRH・LHサージを含む生殖機能に必要不可欠であることが示唆されています。マウスのSCNは約2万個の神経細胞(ニューロン)で構成されており、これらのニューロンは情報伝達に際して様々な神経ペプチド・伝達物質を利用しています。そのため、どのタイプのニューロンがどのようなメカニズムで雌性生殖機能を時間的にコントロールしているかについては未だ不明な点が多いです。 そこで私たちFemale reproductionチームでは、「雌性生殖機能に必要な時刻情報が概日時計中枢であるSCNでどのように生成され・伝達されているか」について探求しています。具体的には以下の3つのテーマについて研究を進めています。
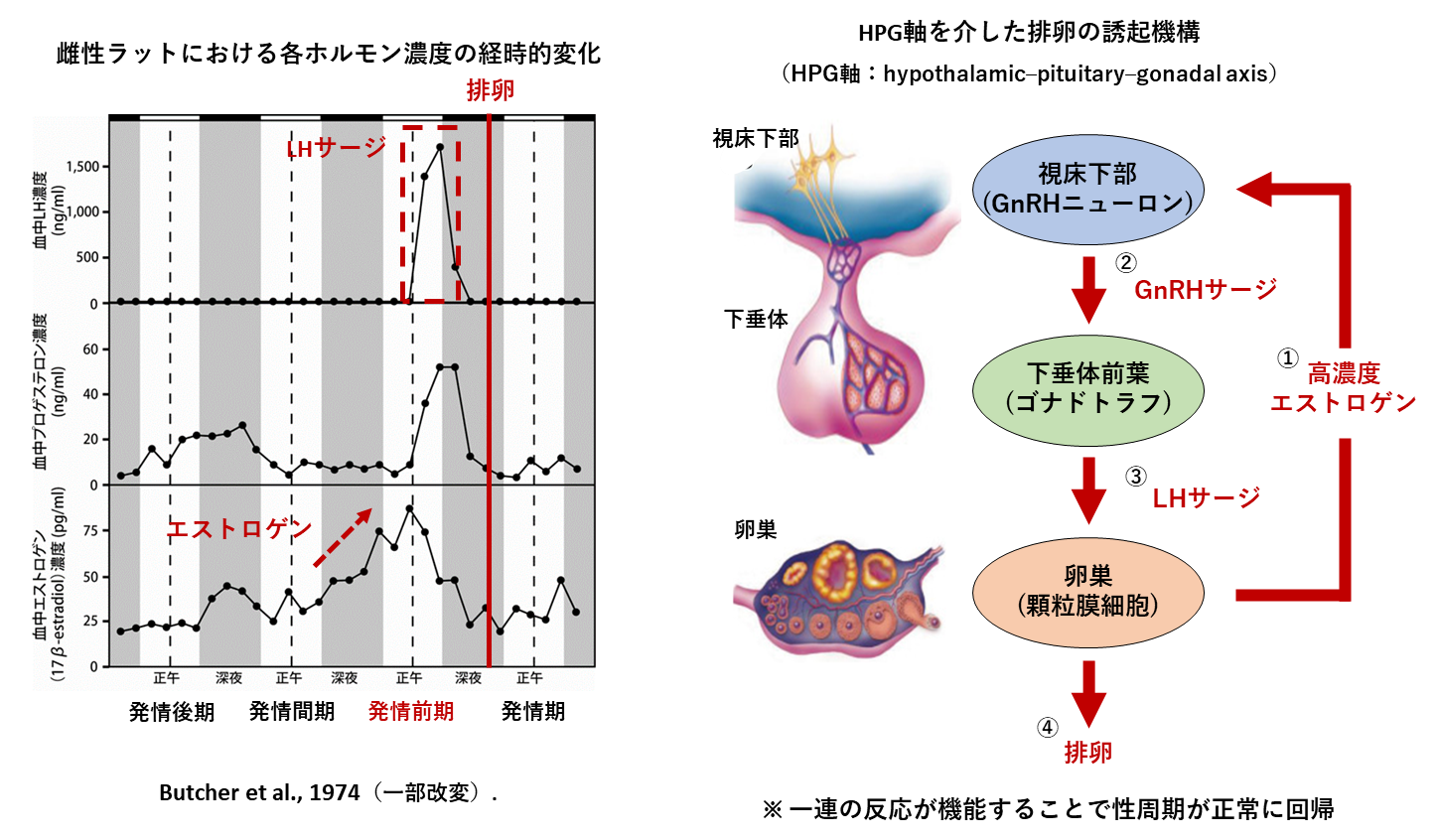
図1 げっ歯類の性周期および排卵のメカニズム
雌性ラットの性周期におけるホルモンの経時的変化(左)および基本的な排卵の誘導メカニズム(右)。
雌性生殖機能における視交叉上核AVP・VIPニューロンの役割
SCNでは様々な神経ペプチド・伝達物質が発現していますが、その中でもSCNの背側部にはアルギニンバソプレッシン(arginine vasopressin, AVP)、腹側部には血管作動性腸管ペプチド(vasoactive intestinal peptide, VIP)を発現するニューロンがそれぞれ局在しており、SCNの主要な神経ペプチドとして機能しています(図2)。近年、解剖学および薬理学的検討により、AVP・VIPニューロンのGnRH・LHサージ機構への寄与が示唆されています。しかし、これらの中には矛盾した結果も多くあり、また、In vivo実験系の研究報告やSCN特異的に操作を行った研究例は少なく、生体内での各ニューロンの機能や役割は未だ不明なままです。そこで本研究では、アデノ随伴ウイルスベクターを用いてAVP・VIPニューロン特異的に遺伝子改変を加えた雌性マウスのLHサージや性周期を観察することで、これらのニューロンの雌性生殖機能における役割・機能を改めて検討しています。 これまでに、シナプスでの神経伝達に関与しているγ-アミノ酪酸(gamma-aminobutyric acid, GABA)の小胞トランスポーター(vesicular GABA transporter, VGAT)をSCNのAVPニューロン特異的に欠損した雌性マウスにおいて性周期が乱れる事を発見しました(図3)。また、ヒト-ジフテリア毒素をSCNのVIPニューロン特異的に発現させた雌性マウスにおいて性周期が乱れる事を確認しています。
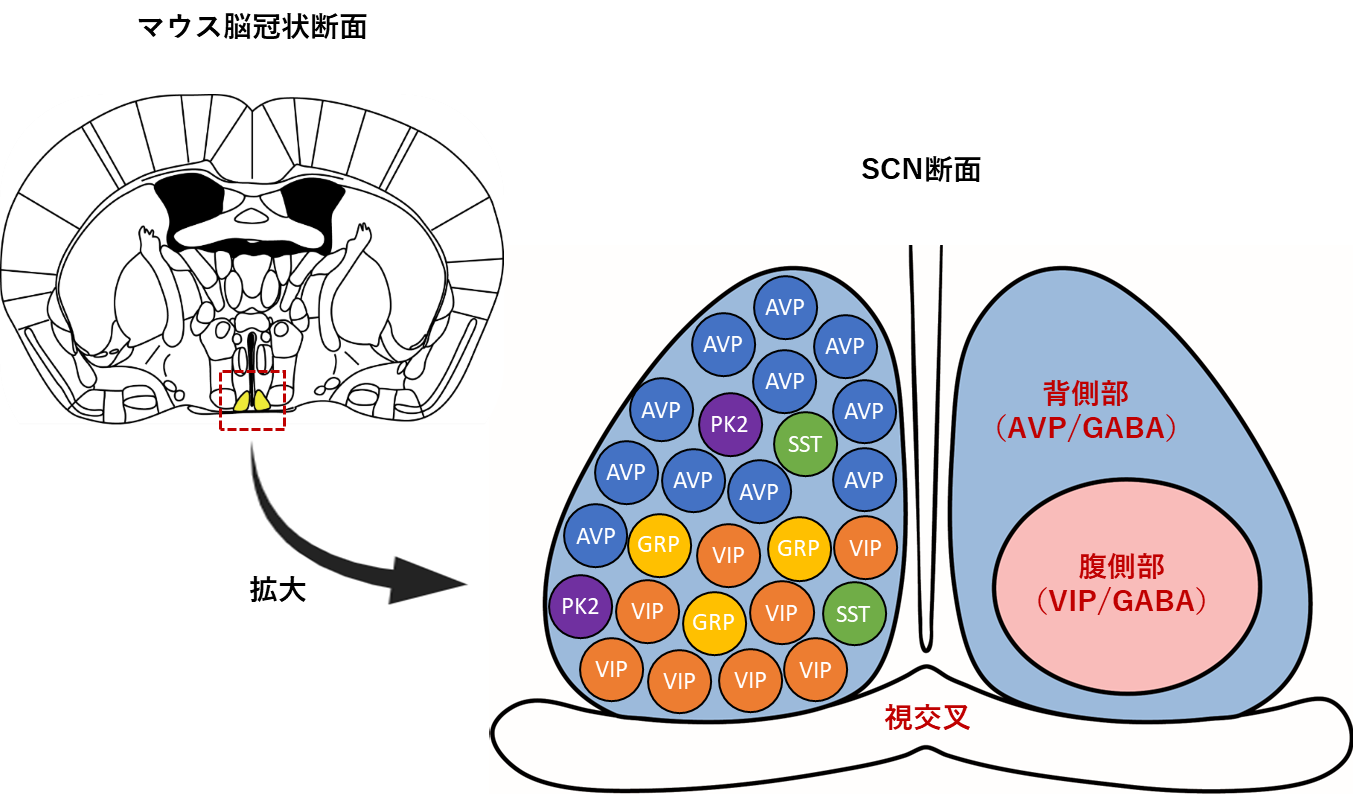
図2 マウスにおけるSCNの解剖学的構造
SCNを含むマウスの冠状断面図(左)および解剖学的なSCNの概略図(右)。